



都说单抗看靶点,双抗看平台,今天这篇文章我们系统的介绍一下我国目前的双特异抗体平台。在前几年,我国的双特异抗体平台在数量上与国外相比大大落后,但是随着近两年的快速发展,国内多数企业都建立了自己独具特色的双特异抗体平台。这些平台既有类IgG平台,也有不含有Fc的片段型平台。构建技术既有Fab融合,纳米抗体融合,scFv融合,也有带电氨基酸突变和二硫键改造,可谓八仙过海,各显神通。据大脸猫统计,目前已经有近30种双抗平台,以下是各个平台的大致介绍(文末有表格汇总)
YBODY
该双特异抗体技术平台由武汉友芝友公司开发,该技术平台双特异抗体包含三个多肽链:一个重链(HC),一个轻链(LC)和一个单链抗体(SC)。这三条多肽链组成的双特异抗体包含三个部分。第一个部分是Fab,该部分主要靶向肿瘤抗原(TAA),第二部分为单链抗体部分(scFv),该部分主要靶向与免疫相关的靶点(IAA),第三部分是由两个不同的抗体CH3组成的Fc。在该技术平台中,抗体的两条CH3部分引入Knob-In-Holes(KiHs)和电荷相互作用,以此来防止同源二聚体的形成。另外,为了防止轻链与重链的错配,其中一个抗体靶点采用单链抗体(scFv).
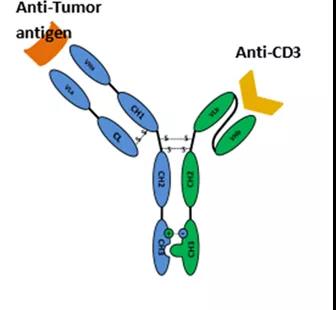
(F(ab)2-(Fv)2-Fc
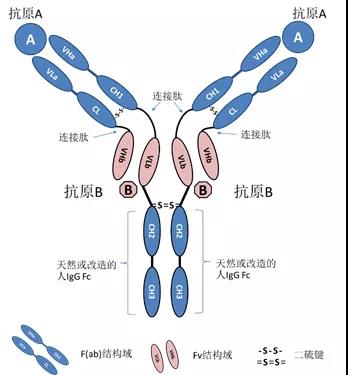
(F(ab)2-(Fv)2-Fc同样由武汉友芝友公司开发,其为四价对称双特异性抗体平台技术,具体为带有完整Fc的对称结构抗体,包括两条相同的重链和两条相同的轻链,轻-重链以及重链-重链之间通过二硫键连接,两个F(ab)结构域结合抗原A,两个Fv结构域结合抗原B,F(ab)和Fv通过连接肽连接。该结构的双抗体可保持与母单抗一致的亲和力,通过该平台技术,友芝友生物已开发一系列双特异性抗体产品。
CRIBTM
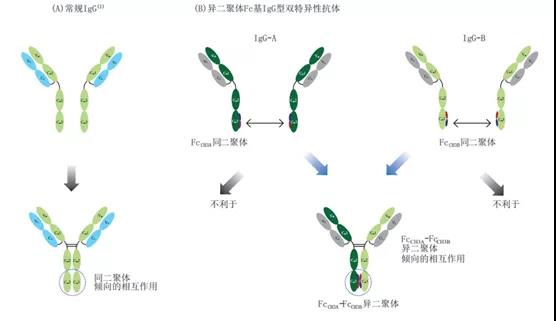
CRIBTM(Charge Repulsion Improved Bispecific):该技术平台由康宁杰瑞开发,该双特异抗体技术平台主要是对抗体的Fc进行改造,使两个不同抗体的CH3部分带相反的电荷,从而防止同源二聚体副产物形成。
ITabTM
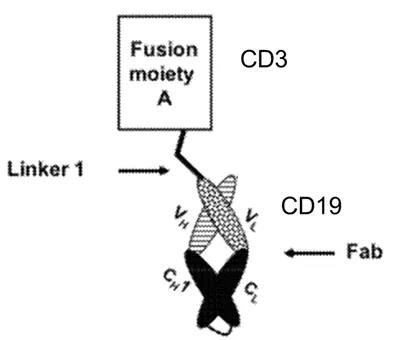
ITabTM(Immune-therapy antibody):该双特异抗体平台由上海键能隆医药公司开发,主要用来激活T细胞,该双抗平台不仅结合CD3激活T细胞,还可以结合肿瘤细胞表面的靶点(图1D)。ITabTM平台以Fab为基础,在Fab的重链和轻链的N端通过linker连接单链抗体(scFv)。
FIT-IgTM
FIT-IgTM(Tetravalent Fabs-In-Tandem immunoglobulins):该双特异抗体由岸迈生物开发,将两个抗体融合到一个抗体平台中—在完整抗体的基础上融合第二个抗体的Fab,为了防止轻链与重链的错配,第二个抗体的轻链融合在第一个抗体的重链的N端,因此该平台中含有三个多肽链,第一个抗体的轻链,第一个抗体的重链和第二个抗体的轻链,第二个抗体的Fab部分的重链部分(VH+CH1)
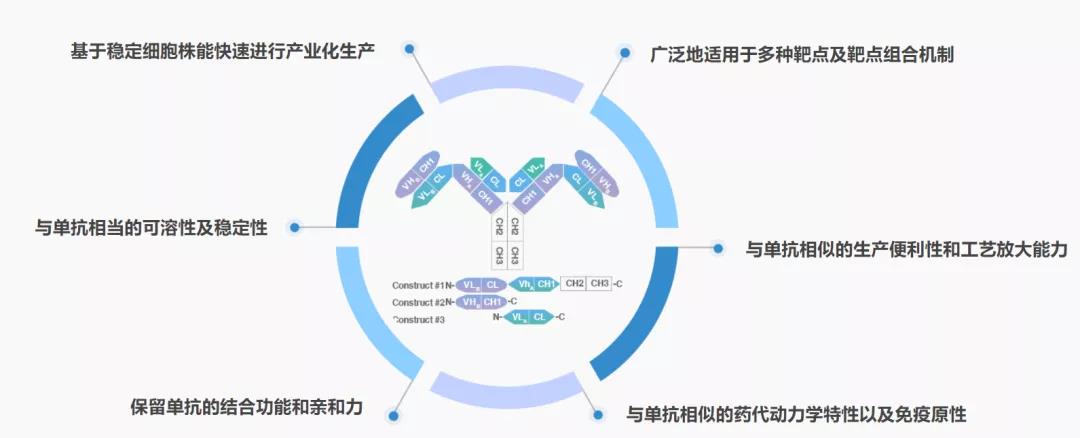
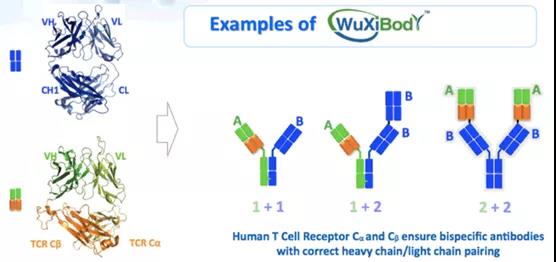
WuXiBodyTM
WuXiBodyTM:该双特异抗体平台由药明生物开发,其主要是借用人T细胞受体(TCR)的恒定去对抗体进行改造,为了防止双特异抗体轻链和重链的错配,将其中一个抗体的LC和CH1用TCR的恒定区α(Cα)和TCR的恒定区β(Cβ)进行替换。该双特异抗体平台比较灵活,即可以形成1+1非对称的双抗,也可以形成2+1非对称形式的双抗(图1G),还可以形成2+2对称形式的双特异抗体。
SMABTM
SMABTM:该双特异抗体平台由金斯瑞生物科技公司开发,其主要是在单抗的基础上融合可以结合第二个靶点的单域抗体(SDA又成为纳米抗体)。因为纳米抗体较小,因此有较高的灵活性,纳米抗体可以融合到单克隆抗体的重链或者轻链的N端和C端,因此可以形成多种结构

恒瑞HOT-Ig双抗
恒瑞医药的双特异抗体采用了药明生物类似的策略,用体内相关异源二聚体蛋白对其中一个抗体的CH1和CL进行替换,从而防止双特异抗体中重链和轻链的错配。不同的是恒瑞医药采用的是Titin(肌联蛋白)和Obscurin(遮蔽蛋白)
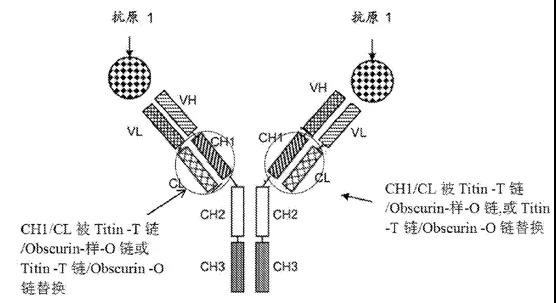
WO2021139758
启愈生物IL-15/IL-15Ra双抗
启愈生物的双特异抗体平台采用了和恒瑞医药及药明生物相同的策略,其用IL-15/IL-15Ra的序列对其中一个抗体的CH1和CL进行替换。利用IL-15/IL-15Ra的高亲和力防止抗体重链和轻链的错配。在防止重链错配方面其采用KIH突变,同时其中一条重链可能还搭配了H435RH435R/Y436F突变,用以消除其与Protein A的结合能力,从而方便后续纯化。

CN113135996
荣昌生物的Hibody双抗平台
Hibody(Hinge-insersion bispecific antibody)结构如下,其是在抗体重链的铰链区插入可以结合第二个靶点的片段,从而使得抗体能够同时结合两个不同的靶点。从其专利来看,第二结合域重点为受体或者配体片段,所以抗体理论上为对称型结构。目前,依据该双特异抗体平台,荣昌生物已经有三款双抗抗体处于准备申报阶段。
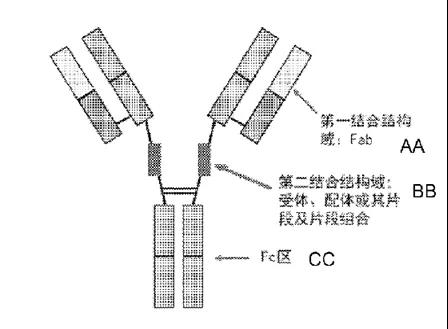
WO2021185337
普米斯生物双特异/多特异抗体平台
普米斯生物的双特异抗体平台包含两种不同的形式:不含有Fc的片断型双特异/多特异抗体平台和含有Fc的双特异抗体平台。
其中不含有Fc的双特异/多特异抗体平台的主体主体结构为Fab,在此基础上融合靶向不同靶点的纳米抗体,其中包含抗HSA的纳米抗体,用以增加抗体的半衰期。另外,其同时在Fab重链或者轻链的末端融合结合其它抗原的纳米抗体,从而形成能同时结合三个不同靶点的抗体。

不含Fc的抗体平台(CN113461824)
而对于含有Fc的双特异/多特异抗体平台,其是在抗体的重链和轻链的N末端融合靶向不同靶点的纳米抗体,该抗体平台的结构为对称型的双特异抗体平台,因此后续生产或者纯化工艺理论上和单克隆抗体类似。
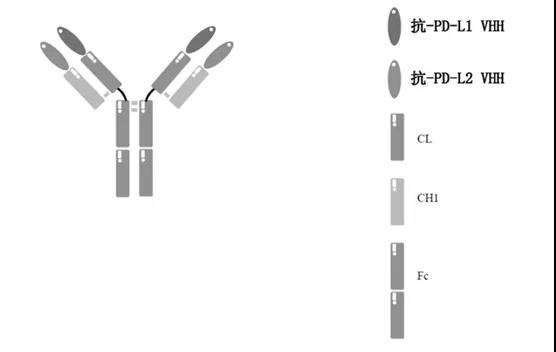
含Fc的双抗平台(CN113461824)
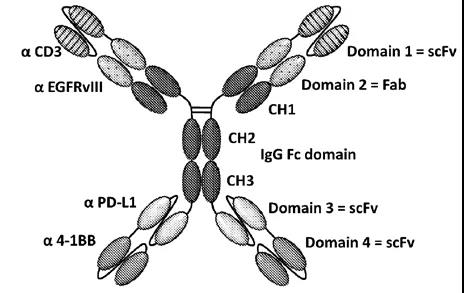
百利药业四抗
百利药业的四特异抗体平台是以完整的IgG抗体为基础,然后在抗体的N末端和C末端融合响应的scFv,该抗体平台从分子量上来看是目前双抗平台中最大的一个,而且抗体中含有多个scFv,因此后期工艺的开发可能也比较具有挑战。
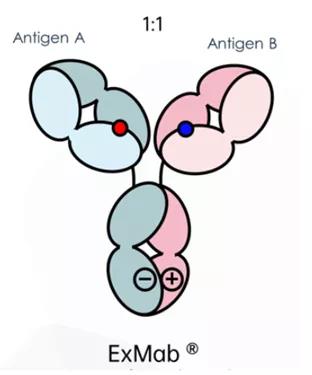
爱思迈ExMab®
爱思迈双特异性抗体平台ExMab®,主要是通过带点氨基酸突变对抗体的Fc进行改造,从而防止抗体重链和重链错配形成Homedimer,而在防止轻链和重链错配方面,其采用疏水或者电荷改造。另外,根据其后续相关专利披露,Fab同时还采用了二硫键配对改造技术。目前爱思迈已经有两款双抗申报临床。
苏州泽璟生物
苏州泽璟生物的双特异抗体平台主要来自于其子公司Gensun Biopharma,主要包括双特异抗体平台和三特异抗体平台。
其三特异抗体平台有两种构建方案;1)在IgG抗体的N端融合VH和VL,并且采用了和罗氏CrssMab类似的技术,同时在抗体重链的C端融合第三个结构域,可以是受体或者配体,或者是特异性结合的肽等。2)利用scFv替换常规的Fab,即双体抗体的一半是scFv-scFv结构(是由两个靶向不同靶点的scFv串联而成),双抗的另外一半为常规的抗体结构。
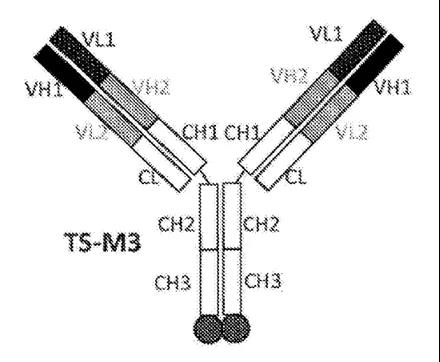
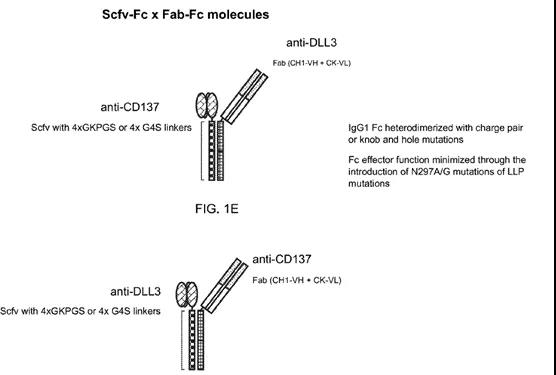
而对于其双特异抗体结构,同样有两种不同的方案:1)采用类似于Ybody的结构;2)类似于安进的延长半衰期的BITE的结构,不同的是,其Fc不是采用串联表达,而是分开表达(理论上应该也加入了KIH突变防止重链的错配)
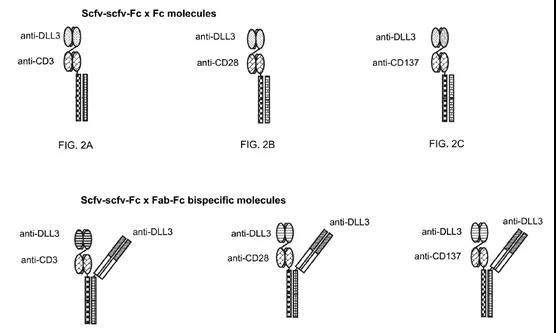
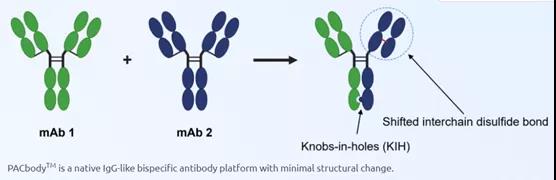
凡恩世PACbody TM 平台
PACbodyTM 平台采用knobs-in-holes技术,在重链CH3结构域上定点突变,促进异二聚体的形成。其次, PACbodyTM 平台通过引入凡恩世自主研发的二硫键位点改造,促进同源轻链和重链的正确搭配(单抗1的轻链和重链通过天然二硫键正确搭配,单抗2的轻链和重链通过改造的二硫键正确搭配)。其在原理上和阿斯利康的DuetMab(原本由MedImmune开发)
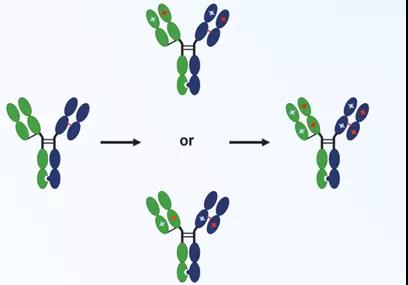
凡恩世SPECpairTM
SPECpair TM(hifted Properties Elicited by Charge pairs)双特异抗体平台的构建采用了带电氨基酸突变,其结构如下图所示,用Knob-In-Hole的结构防止重链错配形成homedimer,在防止轻链错配方面,对不同的链进行带电氨基酸突变,利用同种电荷相互排斥的原理,使得轻链和本身的重链进行配对组装。
凡恩世ATACCbody™
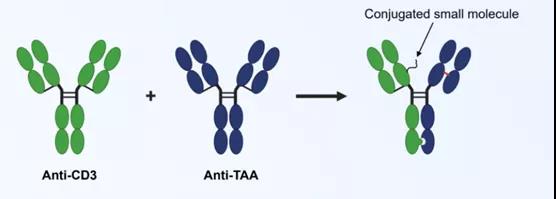
ATACCbody™是在PACbodyTM双特异抗体平台基础上发展而来的新一代肿瘤微环境特异性激活的双特异抗体平台。其原理和Probody类似,不同的是,ATACCbody™是利用小分子(如脂肪酸等)和抗体偶联,利用小分子对抗体的结合表位进行遮蔽,从而使得抗体无法与相应的抗原结合。在肿瘤中,利用肿瘤微环境的特点对小分子进行去除,从而暴露抗体的结合位点,激活抗体的功能并对肿瘤进行杀伤。
和铂HBICETM
HBICETM平台的构建是基于和铂自主开发的HCAb还是转基因小鼠平台,利用小鼠产生的人重链抗体VH或者纳米抗体VHH构建双特异抗体。由于VH或者VHH分子量较小,且为单链,因此在构建双特异抗体时更加方灵活,不会存在传统IgG类双抗的重链和轻链的错配,但是对于的非对称的双抗,其还是需要Knob-In-Hole等技术防止重链和轻链错配。
时迈药业HBiBody和proBibody
HBiBody是由浙江时迈医药开发的双特异抗体平台,其是在Fab的两条链上融合了抗体的Fc,其中利用CH3的突变防止Fc形成同源二聚体,另外在Fab两条链的N端,通过linker融合了可以结合第二个抗原的VH和VL,从而形成了1+1型的双特异抗体。
proBibody和HBiBody一脉相承,不同的是,其对连接第二个抗体VH,VL中的一个linker进行了独特设计,使得修改后的linker可以在肿瘤微环境特异性酶的作用下被切割,从而使得Fab的结合位点暴露并与抗原结合,从而发挥相关作用。
百奥泰双抗
百奥泰的双特异抗体平台为2+2型对称的结构,其在抗体的Fab上融合了可以结合第二个抗原的Fab,为了防止两个不同抗体的轻链和错误配对,其采用了类似于罗氏CrossMab的技术,即将融合抗体的Fab的重链和IgG抗体的轻链融合,而融合Fab的轻链和IgG抗体的重链融合。该技术同样和岸迈生物的FIT-IgTM类似,不同是FIT-IgTM融合的Fab的重链并没有和IgG抗体的轻链连接。
WO2021238932
齐鲁MabPair
MabPair并不是严格意义上的双特异抗体,其为混合抗体,类似于联合用药。其主要是通过在抗体的Fab和Fc上引入带点氨基酸突变,防止在生产过程中两个抗体的重链与重链,或者重链与轻链错误配对,相信介绍见《齐鲁制药 MabPair抗体平台 》
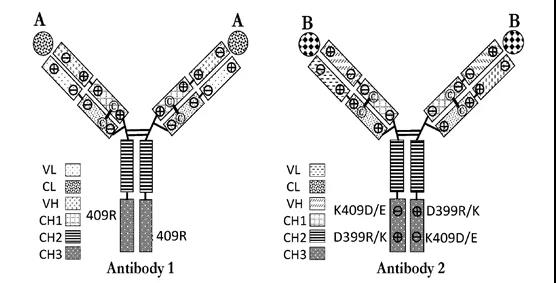
康宁杰瑞开发CRAM平台
CRAM为康宁杰瑞开发的混合抗体平台,与齐鲁的MabPair类似,其同样可以在一个细胞中表达两个不同的抗体。在技术上通过改变电荷对Fc区的CH3域交界面进行修饰,从而建立起有利于Fc同二聚体形成的网络,且阻断不利于Fc异二聚体形成的静电相互作用。在进行共表达时,这些在Fc同二聚体界面上具有电荷异质性修饰的抗体会优先形成且完全保留各成分功能的同二聚体,而不是无活性的异二聚体*产品相关杂质。
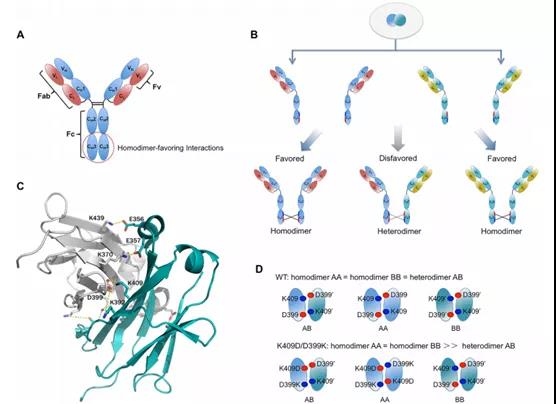
恩沐生物
恩沐生物双/多特异抗体的构建策略是利用两个抗体可变区对其中一个Fab替换,如下图(左)所示,其中H1和L1为第一个抗体的VH和VL,H2和L2为第二个抗体的VH和VL,为了增加抗体VH和VL的正确配对,在H2和L2上进行二硫键突变和正负电荷氨基酸突变。该双/多特异抗体抗体平台在大小上和常规IgG抗体相差不大,但是可以形成2+1或者1+1+1型的多价抗体(下图右,右侧为正常的Fab)。

总结
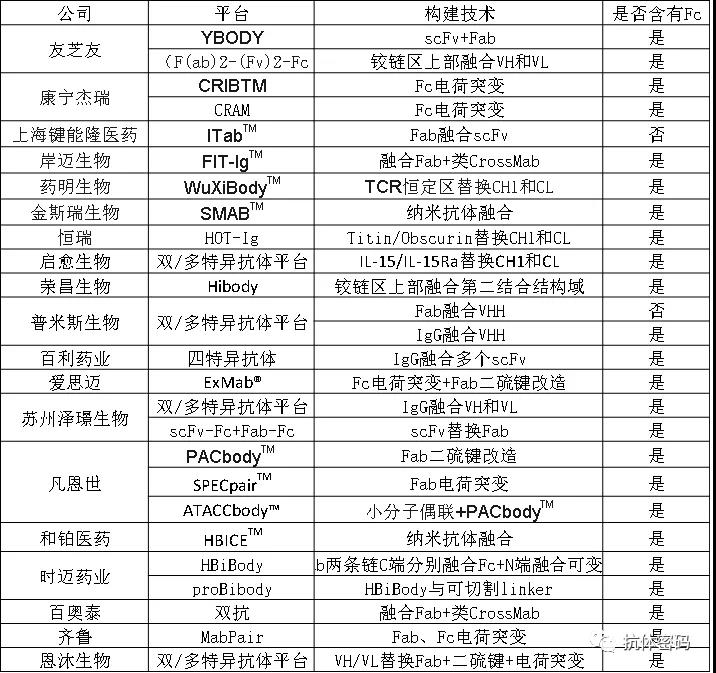
本文我们介绍了26款双特异抗体平台(可能还有一些没有披露或者大脸猫没有了解到,后续会不断更新),这些平台利用不同的技术进行构建:Fab融合,scFv融合,VHH融合,VH/VL替换Fab,电荷改造,二硫键改造,CH1/CL替换等,各具特色。在抗体类型方面,以含有Fc的抗体平台居多,因为Fc可以保证抗体在体内有较长的半衰期。不含有Fc的平台仅有健能隆的ITabTM和普米斯生物的Fab融合VHH的平台,其中普米斯生物通过融合抗HAS抗体延长半衰期。
都说单抗看靶点,双抗抗平台,但是个人认为,平台只是基础,有了好的平台并不能一本万利,后续如何有效的利用和高效的开发才是关键。
来源:抗体密码
声明:本平台注明来源的稿件均为转载,仅用于分享,不代表平台立场,如涉及版权等问题,请尽快联系我们,我们第一时间更正,谢谢!


